








Alpha-1 antitrypsin deficiency (AATD) is an underdiagnosed hereditary condition that promotes the development of lung and liver diseases, and the most common potentially life-threatening genetic condition in Caucasian adults. In this study, the clinical and genetic profile of pulmonary patients from a single center in La Palma Island (Canary Islands, Spain) was assessed to predict how to increase AATD diagnosis.
MethodsAATD was tested in 1,493 pulmonary outpatients without regard to respiratory symptoms and 465 newborns. Variants of the SERPINA1 gene were characterised by real-time PCR, DNA sequencing, molecular haplotyping and phenotyping (AAT isoelectric focusing). Different respiratory pathologies were diagnosed in patients and their levels of serum AAT were measured by nephelometry.
ResultsThe prevalence of pneumological patients with AATD alleles was 30.5%, including PI*S, PI*Z and 6 rare genetic variants. Certain deficiency genotypes were unevenly distributed among patients diagnosed with respiratory diseases: PI*ZZ (71.4%) and PI*SS (34.8%) genotypes were more represented in patients with chronic obstructive pulmonary disease (COPD), whereas PI*MZ (27.7%) and PI*SZ (34.5%) genotypes were more abundant in patients with bronchial asthma. The estimated frequency of PI*S and PI*Z alleles in the general population was 8.2% and 2.1%, respectively. A very significant enrichment (p< 0.01) of PI*S allele, independent of the PI*Z allele, was detected in the clinical population.
ConclusionsAATD diagnosis would improve if both the COPD and the asthmatic patients were included to screening programs. The prevalence of PI*ZZ genotype in La Palma (1/2,162) was relatively high within Spain (average 1/3,344).
Human alpha-1 antitrypsin (AAT) is a glycoprotein with antiprotease and immunomodulatory activity. It is synthesized mainly by hepatocytes and secreted into the bloodstream, where it reaches baseline concentrations of 0.9-1.75 g/L. AAT level in plasma can increase by up to 100% in response to inflammatory or infectious stimuli. The main function of AAT is to inhibit the excess of elastase 2 and proteinase 3 released by activated neutrophils, thus avoiding excessive proteolytic degradation of elastin and type IV collagen in the lungs.1,2 Certain mutations in the SERPINA1 gene (Serine Protease Inhibitor, group A, member 1) cause AAT deficiency (AATD), an inherited condition that predisposes to the development of different diseases, typically pulmonary emphysema and several liver pathologies. While liver disease is triggered by intracellular accumulation of AAT polymers, emphysema is caused by insufficient serum AAT concentration to protect connective lung tissue from the harmful effects of neutrophil proteases.1,2
Hundreds of variants of the SERPINA1 gene have been identified by molecular analysis and about 70 of them have been associated with clinical manifestations.3,4 Normal alleles are called PI*M, with the subtype PI*M1-Val213 being the most common in Europe and North America (44-49%).5 The most common deficient alleles in populations of European ancestry are PI*S and PI*Z, with frequencies of 5-10% and 1-3%, respectively. Nearly 100 percent of the clinical cases of AATD-associated pathologies involve the PI*Z allele, normally as PI*ZZ homozygous or less frequently as compound heterozygous.1
Despite being one of the most common genetic disorders among Caucasians, it is estimated that approximately 90% of individuals with severe AATD remain undiagnosed. In detected cases there has been an average delay of 6 years from the onset of symptoms, often when lung damage is irreversible. Delay in prescribing specific treatment leads to an increase in mortality.6-9 The underdiagnosis can be ascribed to several causes: first, there is unawareness of AATD among physicians; second, the clinical symptoms caused by AATD are essentially indistinguishable from those of other common respiratory pathologies; and third, practitioners request the specific test for AATD diagnosis only if the patient closely matches the classic clinical description of AATD-associated lung disease.2
To analyse, in a local model, how actions on these causes could improve the AATD diagnosis, in the present study the variants of the SERPINA1 gene were surveyed in pneumology outpatients and this information was used to predict what would have been the success of detection of AATD by applying different clinical criteria. In addition, we compare the frequencies of PI*S and PI*Z variants in clinical population with those in the general population to which they belong, to assess whether carriers of one of these deficiency alleles have higher probability of developing respiratory symptoms.
MethodsStudy populationsTwo subsets of subjects from La Palma Island (Canary Islands; Spain) were analysed in the context of AATD: one from a clinical population and one from the general population. For the first subset, pneumology outpatients were recruited during 54 consecutive months (January 2011 to June 2015), from the General Hospital of La Palma, regardless of the cause of the visit or of their respiratory symptomatology. Blood samples for the determination of AAT levels and phenotype or for SERPINA1 gene genotyping were collected in non-anticoagulated tubes or dried blood spots (DBS), respectively. For the second subset, DBS cards were collected from all individuals born in 2014 in the Obstetrics Unit of the same hospital, taking advantage of blood sampling to rule out congenital metabolic diseases. This set of neonates was assumed to be a random and representative sample of the general population from La Palma, whose census in 2014 was 83,456 inhabitants (Spanish National Institute of Statistics).
The study was approved by the hospital's Ethics Committee with the number/date HGLaPalma_2010_7/ September 26, 2010 (patients) and HGLa Palma_2013_12/November 7, 2013 (newborns). Signed informed consent was collected from all patients and the parents of infants. Data on the genotype of newborns were incorporated into the corresponding clinical records and only communicated when requested by the parents. In these cases, and when appropriate, counselling about preventive health behaviours was offered.
Diagnosis of lung diseasesThe respiratory pathologies diagnosed in the pneumology outpatients, following the criteria of specific guidelines, were bronchial asthma, chronic obstructive pulmonary disease (COPD), sleep apnea-hypopnea syndrome (SAHS), hypoventilation-obesity syndrome (HOS) and non-specific bronchial hyperreactivity (BHR).10-13
Genetic characterization of AATD variantsThe approach followed for the genetic characterization of SERPINA1 is summarized in Fig. 1. Genomic DNA extraction was performed in all subjects following an alkaline lysis method from peripheral blood samples deposited on filter paper (Whatman™ 903) and dried at room temperature.14 From the nucleotide sequence of the SERPINA1 gene (NC_000014) amplification primers and HybProbe® fluorescent probes (Supplementary Table 1) were designed to detect, using the LightCycler 480 (Roche) platform, four different mutations: S (rs17580; c.863A>T; p.Glu264Val); Z (rs28929474; c.1096G>A; p.Glu342Lys); ΔPhe52 (rs775982338; c.226_228del; p.Phe52del); and T6 (rs763023697; c.1130insT; p.Leu353PhefsX24). These oligonucleotides were used for real-time PCR and melting analysis in the conditions described elsewhere,15 with the exception of amplification primers for non-S/S and non-Z/Z variants that were used at an annealing temperature of 56°C. Genotyping assays were validated with a set of reference samples previously characterised by DNA sequencing, achieving a good resolution of alleles (Supplementary Figures 1 and 2).
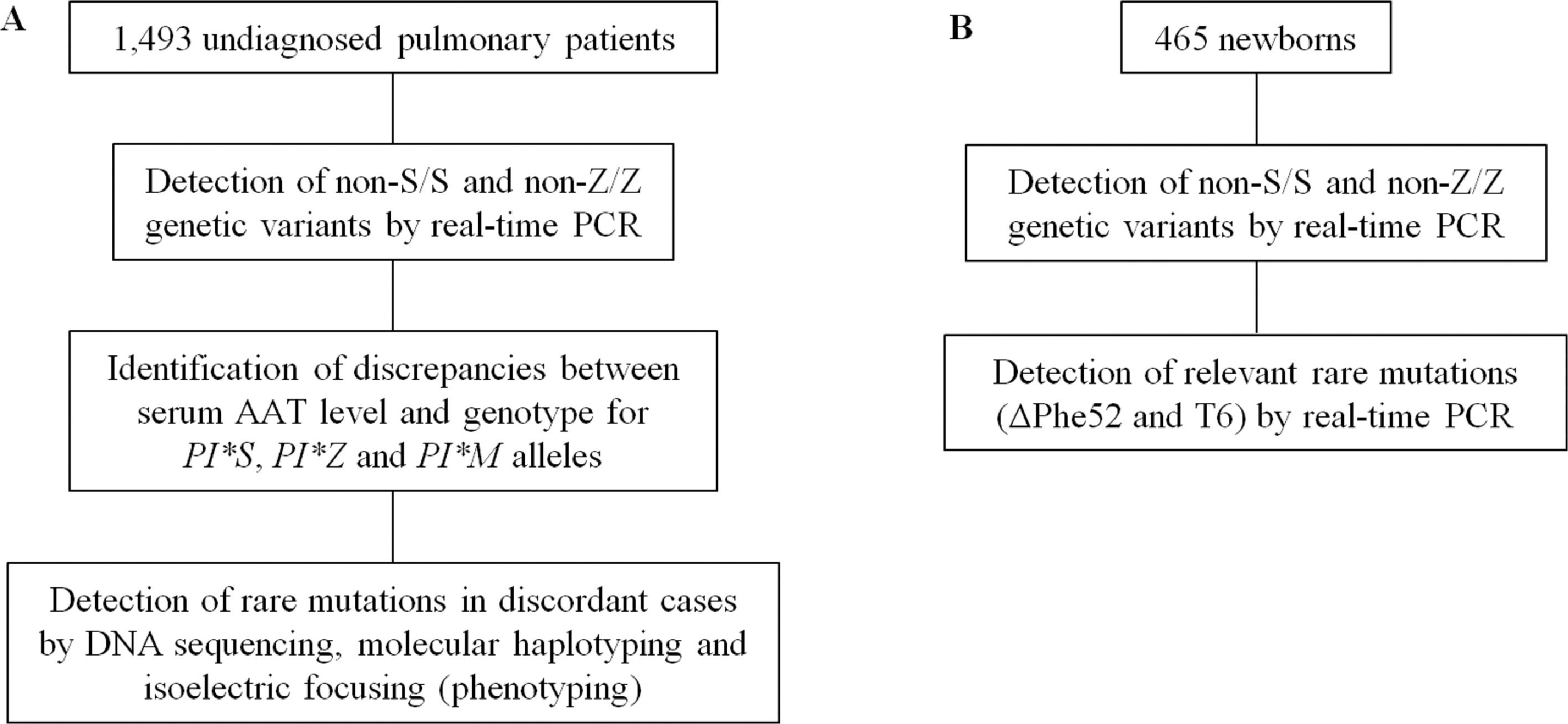
For each patient, serum AAT concentration was determined by immunonephelometry and compared to the reference ranges (95% interval) described for the different combinations of the non-S/S and non-Z/Z genetic variants,16 i.e. the PI*MM, PI*MS, PI*MZ, PI*SS, PI*SZ, and PI*ZZ genotypes (being PI*M = non-S, non-Z; PI*S = S, no-Z; PI*Z = no-S, Z). When the serum AAT level of a patient was less than the lower limit of the corresponding range, genotyping of the SERPINA1 gene was expanded by sequencing the entire coding region and the exon/intron boundaries, using the primers and PCR conditions described elsewhere.17 The nucleotide sequences obtained were compared to the PI*M1-Val213 allele of the SERPINA1 gene (NG_008290.1) to detect variants. In some cases, the complete genetic characterization also required molecular haplotyping by allele-specific PCR and sequencing of the resulting amplicons,15,18,19 sometimes supplementing the diagnosis with data on the AAT phenotype determined by isoelectric focusing and immunodetection.20
Frequencies of alleles and genotypes, and statistical analysisRelative allelic frequencies were determined by dividing the total number of alleles found of each type (PI*M, PI*S, and PI*Z) by the total alleles of the population, expressed as percent. The Chi-Square Test (χ2) was used to determine the goodness of fit between the frequencies of genotypes observed in the general population of La Palma and the expected frequencies assuming Hardy-Weinberg equilibrium. The Hardy-Weinberg principle was applied to calculate the prevalence in the general population of genotypes carrying the defective alleles PI*S or PI*Z.21 The χ2 independence test was used to estimate the significance of the observed differences in the proportion of genotypes between the general population and the clinical population.
ResultsFrequency of PI*S and PI*Z alleles and respiratory pathologies in the clinical populationBlood samples were collected from a total of 1,493 pneumology outpatients. The average age of subjects was 55 years, with a slight predominance of males (56%) and a high frequency of active smokers (20.5%) or ex-smokers (35.7%). The screening for PI*S and PI*Z alleles by real-time PCR in the set of 1,493 patients revealed that 29.7% of subjects were carriers of some of these deficiency allele (Table 1), most of them (87.5%) with PI*MS or PI*MZ genotype. The respiratory disease most frequently diagnosed in this cohort was bronchial asthma (32.5%), followed by COPD (26.9%), SAHS/HOS (11.9%) and BHR (6.4%).
Description of the clinical populations from La Palma Island in terms of diagnosed respiratory diseases and genotypes formed by combination of PI*M, PI*S and PI*Z alleles.
BA: Bronchial asthma; COPD: chronic obstructive pulmonary disease; SAHS: sleep apnea-hypopnea syndrome; HOS: hypoventilation-obesity syndrome; BHR: non-specific bronchial hyperreactivity.
The proportions of patients that were found in each of the groups defined by the variables "lung disease" or “genotype" are shown in Table 1. The group of patients diagnosed with SAHS/HOS presented the lowest incidence of deficiency genotypes (19.6%). The frequency of PI*MS patients was similar in the different respiratory disease groups (15.0-17.2%), but this was not the case for the other deficiency genotypes. PI*ZZ genotype showed the most skewed distribution, since it was exclusively detected in the COPD group, even with a frequency (1.2%) higher than the PI*SZ genotype (0.5%), although only 5 of 7 PI*ZZ subjects detected in the patient cohort (71.4%) have developed COPD. The frequency of PI*MZ and PI*SZ patients was higher in the bronchial asthma (6.3% and 1.9%) and BHR (5.1% and 2.1%) groups than in the COPD group (4.6% and 0.5%). However, because of the differences in the relative frequency of the different lung diseases in the clinical population, the fractions of PI*MZ and PI*SZ subjects from the clinical population that were detected in the bronchial asthma group (27.7% and 34.5%) were much higher than in the BHR group (4.4 and 7.7%). Finally, the frequency of PI*SS patients was higher in the COPD (1.9%) and BHR (2.0%) groups than in the BA group (1.0%), although the fraction of all PI*SS patients that were detected in the COPD group (34.8%) was much higher than in the BHR group (8.8%).
Estimated prevalence of PI*S and PI*Z alleles in the general populationBlood samples were collected from a total of 465 newborns, which represented all municipalities of La Palma in proportion to their number of inhabitants. Table 2 shows their distribution among the six genotypic classes that can be made up of PI*M, PI*S, and PI*Z alleles, which were detected by real-time PCR. Subjects from the general population carrying deficiency alleles were 19.4%. From these data, the estimated frequencies of the PI*S and PI*Z alleles in the general population of La Palma were 8.2% (95% CI: 6.5-10.2) and 2.1% (95% CI: 1.3-3.4), respectively.
Description of the general population from La Palma Island in terms of genotypes formed by combinations of PI*M, PI*S or PI*Z alleles.
| Genotype | Observed number of individuals | Percentage of individuals | Estimated prevalence 1/X (CI 95%) | a Estimated number of individuals (CI 95%) |
|---|---|---|---|---|
| PI*MM | 375 | 80.6 | — | 67,083(63,882-69,871) |
| PI*MS | 67 | 14.4 | 7(5-9) | 11.943(9,312- 15,166) |
| PI*MZ | 17 | 3.7 | 26(16-42) | 3.143(1,931- 5,016) |
| PI*SS | 3 | 0.6 | 150(97-234) | 544(348-843) |
| PI*SZ | 3 | 0.6 | 285(146-565) | 286(144- 557) |
| PI*ZZ | 0 | 0.0 | 2.162(884-5,455) | 38(15-92) |
CI: confidence interval
No significant differences were found between the number of individuals observed within each genotypic class (Table 2) and those expected under the Hardy-Weinberg law (χ2 = 1.431; p > 0.69). Assuming that general population from La Palma is in Hardy-Weinberg equilibrium, it can be predicted the prevalence of the different genotypes composed of the deficiency PI*S and PI*Z alleles, and the number of subjects within each genotypic class (Table 2).
Risk of respiratory disease and PI*MZ and PI*MS genotypesIf carrying only one PI*S or PI*Z allele represents a genetic risk factor for suffering some type of lung condition, the proportion of PI*MM vs. PI*MS or PI*MM vs. PI*MZ individuals would be expected to be significantly different between the clinical population and the general population from which the patients come. The χ2 independence test revealed that this difference was very significant in the case of the PI*S allele (χ2 = 7.05; p < 0.01) and highly significant for the PI*Z allele (χ2 = 10.33; p < 0.0025). Interestingly, like PI*MM homozygous, asthma was more frequent than COPD among PI*MS and PI*MZ heterozygous patients, whereas the opposite was observed in PI*SS and PI*ZZ homozygous (Table 1).
Spectrum of rare AATD variantsPatients whose serum AAT levels did not agree with their genotype for the non-S/S and non-Z/Z variants were analysed in more detail by DNA sequencing and, eventually, by molecular haplotyping. This allowed the detection of 6 rare AATD alleles (Table 3).15,17,22-26 Most of them were found as compound heterozygous combined with a PI*S or PI*Z allele. This set of rare alleles accounted for 2.1% of the deficiency variants detected in the clinical population that, added to the PI*S and PI*Z alleles, cause the total prevalence of deficiency allele carriers in the clinical population to increase to 30.5%. It is noteworthy that we used AAT levels without attending to the inflammatory status of pulmonary patients. This fact has not affected our results on prevalence of PI*S and PI*Z alleles in patient and newborn cohorts, because all subjects were genotyped for these alleles. However, it may have negatively affected the detection rate of rare mutations in the SERPINA1 gene of the patients, although not significantly because of the uncommon nature of these AATD alleles.
Rare alleles and genotypes of the SERPINA1gene, with their corresponding serum levels of alpha-1 antitrypsin, detected in the clinical population.
| Deficiency allele | Mutation | Genetic background | Patient genotype | AAT level (mg/dL) |
|---|---|---|---|---|
| PI*I | c.187C>T; p.Arg39Cys | M425M1-Val21326M2a | PI*MIPI*SIPI*ZI | 91.173.956.8 |
| PI*Mmalton22 | c.226_228delTTC; p.Phe52del | M2 | PI*ZMmalton | 15.1 |
| PI*Mpalermo23 | c.226_228delTTC; p.Phe52del | M1-Val213 | PI*SMpalermo | 46.3 |
| PI*Q0la palma18 | c.226_228delTTC; p.Phe52delc.863A>T; p.Glu264Val | M1-Val213 | PI*ZQ0la palma | 8.5 |
| PI*Q0ourém15,24 | c.1130insT; p.Leu353PhefsX24 | M3 | PI*MQ0ourémPI*SQ0ourémPI*ZQ0ourém | 76.1b38.614.4 |
| PI*Zla palma17 | c.321C>A; p.Asn83Lys | M1-Val213 | PI*MZla palma | 75.6 |
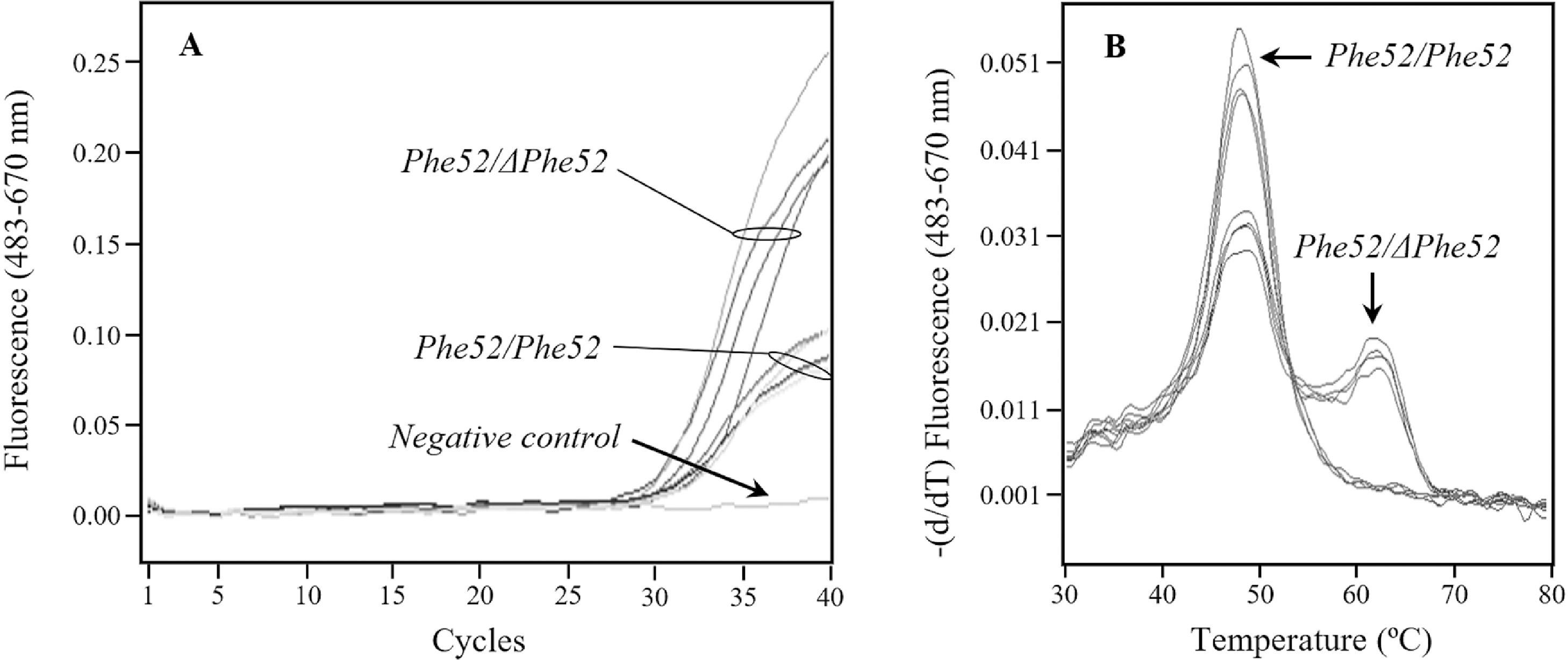
The rare deficiency alleles of the SERPINA1 gene most common in the clinical population from La Palma were the null allele PI*Q0ourém and the PI*I allele, with 5 and 3 carrier patients from a total of 12 cases, respectively. Since there has been an important historical contribution of Portugal to the peopling of La Palma, a search for PI*Q0ourém allele by real-time PCR (T6 variant; supplementary Table 1) was carried out among the 465 neonates, but results were negative. The fact that ΔPhe52 variant were detected in three different genetic backgrounds in the clinical population (PI*Mmalton, PI*Mpalermo and PI*Q0la palma; Table 3) led us to design a specific genotyping assay based on real-time PCR, but the screening of this mutation among the 465 neonates was unsuccessful.
DiscussionThe high frequency of PI*S allele in the population of La Palma (8.2%) is slightly lower than those found in the Iberian Peninsula (11.4% in Portugal and 10.4% in Spain), but similar to those in southwestern France (7.5%) and higher than those found in other countries in the European continent.21 The frequency of PI*Z allele in La Palma (2.1%) is comparable to that found in Portugal (2.1%) and slightly higher than that in Spain (1.7%). In Europe, it is only surpassed by those found in some northern countries as Latvia (4.5%) or Denmark (2.7%).27 The prevalence of the PI*ZZ genotype in La Palma (1/2,162) is relatively high within the Spanish territory (average 1/3,344).28 Therefore, it would be advisable to perform screening in this region aimed at identifying individuals at high risk of diseases associated with AATD.
Our data on SERPINA1 genotypes and respiratory symptomatologies in a set of 1,493 pneumology outpatients could be analysed in search for causes of AATD underdiagnosis, keeping in mind the limitations of the local model represented by La Palma Island. If our genetic analysis of all pulmonary patients would have been focused exclusively on the COPD group, as recommended by some reference guides,29,30 the fraction of subjects identified as carriers of PI*S and/or PI*Z alleles would have been reduced from 29.7% to 6.3%. Although the majority of PI*ZZ patients (71.4%) would be identified following this recommendation, only a small fraction of PI*MZ (17.0%) and PI*SZ (7.7%) patients could be detected, contributing to the underdiagnosis of AATD.2,6,9,31 The inclusion of both asthmatic and COPD patients in our genetic analysis of the clinical population, as recommended by certain reference guides for AATD screening programs,32-34 would significantly increase the success of detection of PI*MZ (44.7%) and PI*SZ (42.2%) patients. It should be taken into account that this observation may be related to local factors that determine the high prevalence of asthma in the Canary Islands.35,36
It is solidly demonstrated that PI*ZZ and, to a lesser extent, PI*SZ genotypes represent a risk factor for the development of dyspnea, cough, early onset pulmonary emphysema and air flow obstruction.2 Our results indicate that the frequency of both PI*MS and PI*MZ genotypes is significantly higher in the pneumology outpatients population than in the reference general population. Although the number of evidence pointing to the PI*MZ genotype as a risk factor for lung conditions is increasing,2,37 the clinical status of PI*MS individuals remains controversial.38 It would be interesting to find out what additional risk factors contribute to the enrichment in PI*MS genotypes that we observed among pneumology outpatient and that was not detected in another similar study, probably because in this case the general population was compared to the results of screening aimed at the detection of AATD.39
ConclusionsAccording to our data, the inclusion of asthmatic patients in AATD screening programs would significantly increase success in detecting this inherited condition.
We thank the health authorities of the island of La Palma for facilitating the realization of this research work, and Grifols S.A. for financial support. Jordi Bozzo and Eugenio Rosado (both from Grifols) are acknowledged for editorial support in the preparation of the manuscript.





{kind=link}
{kind=link}